
Platelet-derived vesicles: diagnostic and predictive value in cardiovascular diseases


Abstract
There is emerging evidence that endogenous microvesicles released by platelets may play a pivotal role in pathogenesis of numerous diseases including stable and unstable ischemic heart disease, atherosclerosis, hypertension, idiopathic pulmonary hypertension, atrial fibrillation/flutter, acute and chronic heart failure, as well as systemic vasculitis, systemic lupus erythematosus, antiphospholipid syndrome, transplant rejection, diabetes-induced angiopathy. Microvesicles that leased from the platelets exist as cargo for a large of number biological-active molecules including regulatory peptides, hormones, growth factors, active molecules coordinating cell-to-cell cooperation. Moreover, platelet-derived microvesicles are concerned about a conspicuous component in regulating angiogenesis, cardiac protection, and vascular repair. Recent animal and clinical studies have shown that the number of circulating platelet-derived microvesicles may sufficiently increase in accelerating atherosclerosis, acute atherothrombotic events, i.e., recurrent ischemia, myocardial infarction, thrombosis, hypertension, and stroke, microvascular inflammation. It is suggested that exaggerating levels of circulating platelet-derived microvesicles is strongly associated with endothelial dysfunction. The review is discussed the role of platelet-derived vesicles in the pathogenesis of the cardiovascular disease, as well as diagnostic and predictive capabilities of platelet-derived vesicles as biomarkers of a natural evolution of the diseases.
Keywords
Introduction
Cardiovascular (CV) diseases are the most common causes of death in the general population worldwide[1]. Moreover, the attributable impact of CV disease on mortality of the general population in both developed and developing countries tends to be steadily raised[2,3]. Although diagnosis and therapy of CV diseases are markedly reported in contemporary clinical statements and guidelines, prediction of CV events and personifying risk stratification for patients with established CV disease remains to be challenged[4,5]. For instance, the presence of several comorbid conditions (diabetes mellitus, abdominal obesity, dyslipidaemia, insulin resistance, rheumatic and autoimmune disease, kidney disease, cognitive impairment, peripheral artery disease, liver and thyroid dysfunctions) sufficiently modifies as prognostication of CV diseases, as well as both clinical management and the therapeutic approach in CV disease. In this context, there is a strong tendency to use integrative complementary therapies of CV disease based on an individual assay of risks and benefits of precise biomarker controlled point-of-care[6]. The identification of therapeutic targets a blockade of which could have a positive impact on CV-related clinical outcomes diminishing a probability of poor prognosis is a promising approach for contemporary strategies in the treatment of CV disease. Consequently, non-invasive circulating biological markers reflecting various stages in the pathogenesis of CV disease can be surrogate indicators for successful diagnostic, stratification and treatment approaches.
Extracellular vesicles (EVs) are small sized (range of diameter is approximately < 1000 nm) particles originated from various cells and deliver biologically active molecules (peptides, RNAs, active molecules, growth factors, hormones, and enzymes) from mother cells to recipient cells through binding, fusion or endocytosis appearing to be a core element in cell-to-cell cooperation[7,8]. They are involved in the pathogenesis of CV diseases through numerous biological mechanisms that support microvascular inflammation, arterial stiffness, vascular calcification, atherosclerotic plaque shaping and rupture, endothelial dysfunction, hypercoagulation and thrombosis, cardiac remodelling, kidney dysfunction[9,10]. Additionally, the EVs play a pivotal role in tissue reparation, angiogenesis, and neovascularization via intracellular signalling cascades[11,12]. In fact, the EVs mediate autocrine and paracrine signals that are enabled to reconstructing the homeostatic micro-environment in heart and vessels[13]. EVs have released a wide spectrum of circulating cells, including endothelial cells, cardiac myocytes, red blood cells, mononuclear/macrophages, other antigen presenting cells, as well as platelets. EVs, which are produced by endothelial precursors and mature endothelial cells, as well as mononuclear, are under investigation for a long time respectively and there is strong evidence regarding the predictive capability of an altered circulating number of EVs in established CV disease[14,15]. On the contrary, the exact role of the platelet-derived microvesicles in the pathogenesis of CV disease remains to be not fully discovered. Controversies regarding the role of platelet-derived vesicles in the CV diagnosis, stratification, and EV-guided therapy mainly affect uncertain methods of identification of immune phenotypes. For instance, there is not strong evidence regarding the platelet-derived vesicles bear numerous protein markers, such as markers of the megakaryocytic lineage (CD110), platelet collagen receptor (CD36), tetraspanins (CD63, CD9 and CD81), P-selectin (CD62), heat shock proteins (HSP70) and integrins (CD41, CD42, CD51, CD61), which are found in other types of the EVs of similar size. Another controversy is associated with storage of whole blood that can lead to releasing the EVs from platelets into the blood sample. Taking together, analytic errors should be paid into consideration prior to an interpretation of the test result for EVs’ measure. Yet, numerous studies have yielded serious limitations in the diagnostic and predictive value of platelet-derived EVs because of small sample size, low sensitivity for clinical end points and none randomized design of the clinical trials[9,10,14]. However, possible diagnostic and discriminative values of the EVs have been still widely debating. The review is discussed: (1) the role of platelet-derived microvesicles in the pathogenesis of CV disease; (2) diagnostic and predictive capabilities of platelet-derived microvesicles as biomarkers of a natural evolution of the diseases.
Extracellular vesicles: definition
EVs are defined as small-sized (< 1000 nm) cell membranous originated structures that secreted by various cells (platelets, red and white blood cells, endothelial cells, precursors/stem cells, cardiomyocytes, and even tumor cells) into human fluids and contained wide range of proteins, hormones, signalling molecules, lipids, mRNA, miRNA, and others[16,17]. There are at least two distinct subpopulations of the EVs [exosomes and microvesicles (MVs)], which distinguish each other in size, compounds, immune phenotypes and wide range of formation modes. Exosomes are sufficiently smaller to MVs (size average is 30-120 nm) and they are shaped by the endocytosis of multivesicular bodies and shed from cells by exocytosis. On the contrary, the MVs (size average is 40-1000 nm) are defined as membrane-bound vesicles, which are released from the plasma membrane. Additionally, surface-specific antigens, which are presented for both populations of EVs, are different, but not specific for types of EVs. They overlap between exosomes and MVs that does not allow differencing both populations of the particles, but the size of particles can be used for differentiation of them. Physiologically, EVs are cargo for molecule transfer, auto/paracrine signaling, cell-to-cell cooperation, playing a pivotal role in immune response, vascular integrity, cellular senescence, tissue reparation and development, angiogenesis and neovascularization[18,19]. In fact, the EVs are able to mediate the exchange of appropriate information between various cells. Finally, the EVs were found witnesses and biomarkers of several diseases, such as CV, autoimmune, infective, metabolic, renal, rheumatic, neurological, ocular diseases and malignancy[20-23].
Platelet-derived vesicles
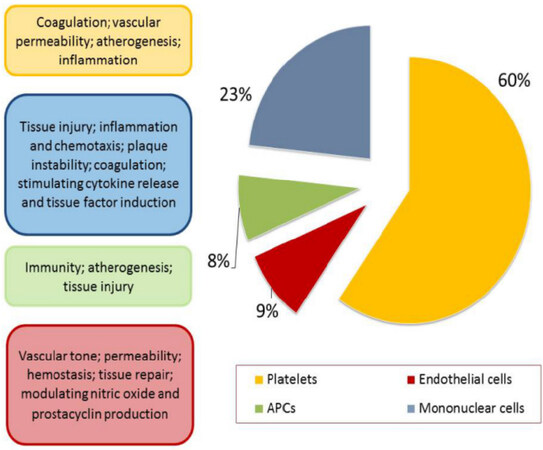
The population of the platelet-derived vesicles is the largest in other types of EVs in the circulation [Figure 1]. Indeed, the majority of the blood-borne EVs are thought to originate from the megakaryocytes, i.e., either from circulating platelets or directly from the platelet precursor cells, which reside in the bone marrow. Although coagulation, thrombosis, vascular senescence and permeability are most common and well-established biological processes, in which platelet-derived vesicles are embedded, the role of them in CV diseases has not to be exhausted a crucial impact on hemostasis. Interestingly, the platelet-derived vesicles display both pro- and anticoagulant properties depending on the presentation of the phosphatidylserine (PS) of the platelet-derived vesicles in the tenase and prothrombinase complexes, as well as an expression of tissue factor on the surface of the platelet-derived vesicles. Both PS and tissue factor are able to initiate the extrinsic coagulation pathway involving active Factor VII. On the other hand, the platelet-derived vesicles were found to have an ability to express glycoprotein Ib and bind the annexin V, protein S and activation of protein C that led to the anticoagulant effect. Thus, the balance between pro-coagulant and anti-coagulant properties of the platelet-derived vesicles plays an important role in supporting of hemostasis[24,25].
Intriguingly, there is evidence that the platelet-derived vesicles are cargo for several molecules, including chromatin, hormones, proteins, which mediate cell-to-cell cooperation, immune reaction, inflammatory response, and reparation[18-22]. It has been found that the main fraction of miRNA is localized in the platelet-derived vesicles, which protect miRNA from degradation in the circulation. Thus, the platelet-derived vesicles are considered a core element of transfer gene-regulatory function from mother cells to target cells[26]. Additionally, the platelet-derived vesicles transfer monomeric C-reactive protein mediating inflammatory reaction from target cells/tissues and activate endothelial cells and their precursors[27]. Therefore, the platelet-derived vesicles may enhance mobbing, migration, aggregation, and accumulation of circulating leukocyte on selectin-expressing substrates mediating increased permeability of the endothelium[28]. Moreover, the platelet-derived vesicles can induce leukocyte to release inflammatory cytokines, such as interleukin (IL)-1β, IL-6, IL-17, interferon-gamma, tumor necrosis factor (TNF)-alpha, monocyte chemoattractant protein-1, and matrix metalloproteinase (MMP)-6, MMP-9, which promote endothelial inflammation, worsening vascular integrity and endothelial dysfunction. There are data clarifying that the platelet-derived vesicles can support differentiation of T cells with regulatory phenotype into T cells with pro-inflammatory phenotype[29]. Although the platelet-derived vesicles can transfer lipoxin A4, which acts as an anti-angiogenic factor for endothelial precursors and mature endothelial cells, yet platelet-derived vesicles can induce angiogenesis and vascular reparation through augmenting endothelial progenitor cell differentiation into peripheral blood mononuclear cells[30,31].
Elevated levels of the platelet-derived vesicles were determined in patients with acute coronary syndrome, acute myocardial infarction, heart failure, arrhythmia, atherosclerosis, as well as heparin-induced thrombocytopenia, thrombotic complications, hemolytic uremic syndrome, while developing of abdominal obesity, diabetes mellitus, antiphospholipid syndrome, and sepsis was not associated with increased circulating level of platelet-derived vesicles[32]. Probably, this evidence may relate to the mechanisms regarding the release of MPs from platelets. It has suggested that in various cases circulating platelets are likely to adhere to leukocytes or endothelial cells at the activation site and that the circulating platelet-derived MPs are likely to be a residue of activated platelets[33].
It has been found that upon activation, blood platelets are able to release two types of EVs, i.e., microparticles and exosomes[34,35]. Platelet-derived microparticles (also referred to microvesicles) are characterized by an average of the size of 70-100 nm and the PS presentation at their surface, which mediates their capabilities in coagulation and thrombosis[36]. On contrary, platelet-derived exosomes are characterized by their small size (< 80 nm) and the presence of CD63 antigen, which is known as the tetraspanin that belongs to the transmembrane 4 superfamily, while larger vesicles (> 0.5 µm) containing chromatin materials are referred to apoptotic bodies[35,36].
Majority of positive CD41 EVs have been reported to be derived from platelets. CD63 plays a pivotal role in the role in the activation of cellular signaling cascades that trigger Akt, FAK-related non-kinase polypeptide/protein tyrosine kinase-2 and mitogen-activated protein kinases, attenuates an adhesion of circulating monocytes/macrophages onto endothelial cells through the regulation of P-selectin (CD62) trafficking, as well as promotes vascular endothelial growth factor receptor (VEGFR) signalling and internationalization[37]. All these factors are involved in cell-to-cell cooperation and cell interactions with the extracellular matrix. Moreover, the EVs expressing Ms4a2/FceRI on the surface are able to be triggers for degranulation of circulating cell in response to other stimuli[37]. Additionally, some EVs may express CD41 (Integrin alpha-IIb) on their surface, which is a complex with CD61 may be a receptor for thrombospondin, fibronectin, vitronectin, fibrinogen, and von Willebrand factor playing a crucial role in blood coagulation[38,39]. The populations of the platelet-derived EVs may distinguish each other in not just their size and immune phenotype, but yet they can have a difference in a density (utilized as sucrose density gradient in ultracentrifugation), proteomic and lipidomic profiles[40]. Labeling of platelet-derived EVs is reported in Table 1.
The baseline characteristics of 312 patients at the time of direct-acting antiviral treatment
| CD marker | Alternative name | Prominent feature | Cell labeling | Expression | Significance |
|---|---|---|---|---|---|
| CD9 | Tetraspanin (Tspan 29) | Key regulator of intracellular signaling, proliferation, activation, survival, migration, invasion, cell adhesion, and diapedesis | Platelets, B cells, CD4+ T cells, CD8+ T cells, natural killer cells, granulocytes, monocytes and macrophages, and immature and mature dendritic cells | Low | Non-specific lymphoid/hematopoietic marker |
| CD23 | Fc epsilon RII (FcεRII) | Interaction with many ligands (IgE, CD21, MHC class II, integrins), exhibiting pleiotropic cytokine-like activities, sustaining the growth of activated B-cells, differentiation of myeloid precursors, thymocytes and bone marrow CD4+ T cells, degranulation of platelets | Platelets, T and B lymphocytes, polymorphonuclear leukocytes, monocytes, follicular dendritic cells, intestinal epithelial cells, bone marrow stromal cells | Low | Non-specific lymphohematopoietic marker, a diagnostic marker for neoplastic and autoimmune inflammatory disease |
| CD31 | PECAM-1, GP IIa, hec7 antigen, EndoCAM | Key inhibitory receptor in circulating platelets and leukocytes, major endothelial mechanosensor, a modulator of vascular permeability, and leukocyte trafficking, and cell-to-cell cooperation, a core player in thrombosis, inflammation and the immune response | Platelets, leukocytes, monocytes/mononuclear, endothelial cells, EPCs | High | A maker of endothelial integrity, vascular permeability, platelet degranulation, endothelialopathies in tumor angiogenesis and the growth and development of hemangiomas |
| CD36 | GP-IV, Scarb3 | Recognition of thrombospondin, fatty acids, collagen, adhesion of platelets | Monocytes, macrophages, platelets, adipose cells, endothelial cells, PECs, epithelial cells | High | Platelet collagen receptor |
| CD41 | αIIb integrin, platelet GPIIb | Key regulator of growth and differentiation | Hematopoietic progenitors in embryo, specific to the megakaryocytic/platelet lineage | Moderate | Markers of all progenitors (megakaryocytic, granulomacrophagic, erythroid and pluripotent cells) |
| CD42 (a, b, c) | Integrin subunit αIIb, GPIIb | Regulator of platelet activation and adhesion | Platelets | High | Marker of platelet aggregation |
| CD49f | GPIc, integrin α6-chain, ITGA6, ITGA6B, VLA 6 alpha Subunit | Regulator of embryogenesis, cell adhesion and cell migration through T-cell receptor/CD3 complex | T cells, endothelial cells, epithelial cell, macrophages, monocytes; platelets and stem cell | High | Marker of blood cell adhesion and T-cell proliferation |
| CD51 | Integrin subunit alpha V, vitronectin receptor | A trigger of cell adhesion and signal transduction, the regulator of angiogenesis and cancer progression | T cells, endothelial cells, platelets | High | Marker of cell adhesion to sequences of von Willebrand factor, vitronectin, fibrinogen, and thrombospondin |
| CD61 | GP3A, integrin subunit beta 3 | Regulator of cell adhesion and cell-surface mediated signaling | Endothelial cells, Macrophage, platelets | High | Marker of human megakaryocytic cell lineages |
| CD84 | Cell surface antigen MAX.3, Ly-9B, SLAMF-5 | Regulator of signaling and adhesion in lymphocytic cells and other cells | Thymus and spleen cells, cells of megakaryocytic lineages | Moderate | Marker of adhesion interactions between T lymphocytes and accessory cells, platelets |
| CD92 | Choline Transporter like Protein 1 | Regulator of cell growth and differentiation | A subset of bone marrow stromal cells, cells of osteogenic, megakaryocytic and adipogenic lineages | High | Marker of human bone marrow stromal cells, and cells of megakaryocytic and adipogenic lineages |
| CD110 | MPL proto-oncogene, TPO-R | Regulator of hemopoiesis | A subset of stem cells and cells of megakaryocytic lineage as well as platelets | High | Marker of megakaryocytic lineage |
| CD147 | BSG, EMMPRIN | A trigger of cell proliferation associated with the TGF-β/Smad4 signalling pathway, cell adhesion, and T-cell activation | Endothelial cell, epithelial cells, lymphocytes, mononuclear, platelets | Low | Marker of cell growth, adhesion, and proliferation, malignancy, angiogenesis, vascular permeability |
| CD151 | GP27, membrane glycoprotein SFA-1, platelet-endothelial tetraspan antigen-3 | Regulator of the VCAM-1 activity during lymphocyte and platelet recruitment | Endothelial cells, platelets, T-cells | High | Marker of fibrosis, vascular inflammation and remodelling, carcinogenesis, vascular permeability, endothelial dysfunction |
| CD226 | DNAM1, PTA1, TLiSA1, DNAX | An adhesion molecule involved in NK and T cell-mediated cytotoxicity | T cells, NK cells, platelets, mononuclear | Low | Triggering cytotoxicity and cytokine secretion by T and NK cells |
| GARP | - | A key regulator of a network between Tregs and its targets | Activated Tregs, mature peripheral naive CD4+ T cells, platelets | Low | Marker of activated Tregs and degraded platelets |
| LAP | - | Phagosome maturation, stabilization of the cargo to prolong antigen presentation on MHC, recruiting LC3 molecules to phagosome membranes class II molecules | Macrophages, dendritic cells, and platelets | Low | A marker of the catabolic process involved during exogenous antigen processing, recognition of apoptotic, necrotic, or entotic cells, degradation of platelets |
Although there are difficulties in identification of distinction between two types of EVs (microparticles and exosomes) in cargo potency, immune heterogeneity and size averaging of EV populations allows identifying them using several methods for their isolation and quantification, including conventional and cryo-enhanced electron microscopy, immuno-gold labelling, laser-scanning confocal microscopy, nanoparticle tracking analysis and flow cytometry[41,42]. However, lack of generally accepted methods as a standard for determination, isolation, and quantification of the platelet-derived EVs, sufficiently limits our efforts in understanding their biological role in the pathogenesis of several states and diseases[43,44]. The review is considered a possible role of both types of EVs (microparticles and exosomes) in CV diseases.
Platelet-derived vesicles in CV disease
There is a large body of evidence regarding the pivotal role of platelet-derived vesicles in the pathogenesis of CV diseases[45-47]. In a result of several triggers, which are reported Figure 2, platelets are activated, aggregated, then they changed their contain enriching phospholipids and GPs, prepare for vesiculation via shaping spaced architectonic and exocytosis and release vesicles into the circulation. Regulation platelet vesiculation is mediated via platelet Ca2+-ATPases by cyclic AMP through the phosphorylation of the Rap1 protein.
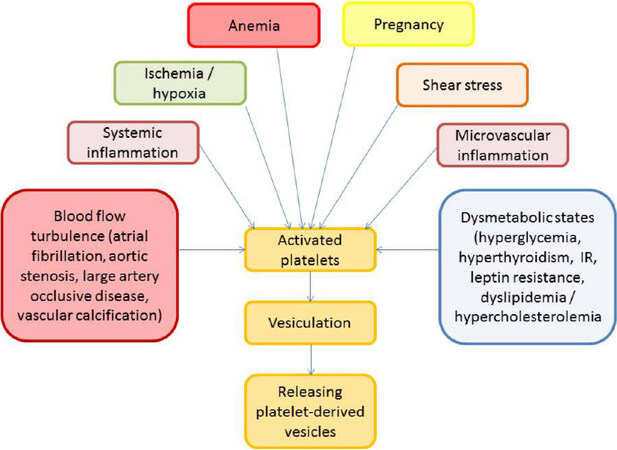
Figure 2. Common triggers for platelet vesiculation. IR: insulin resistance
The platelet-derived vesicles exhibit haemostatic properties, mediate aggregation of blood cells including whole platelets, promote coagulation, stabilize the vasculature and maintain endothelial cell barrier integrity[48]. Therefore, the platelet-derived vesicles may demonstrate pro-atherogenic, pro-inflammatory, and immunomodulatory effects, and even anticoagulant activity depending on triggers that induce their releasing and the expression of their surface certain molecules[49-51]. For instance, phospholipid-enriched platelet-derived vesicles that express Annexin V possess as pro-atherogenic and pro-coagulant particles[52]. On the contrary, Annexin V-negative platelet-derived vesicles were found rather anticoagulant than pro-coagulant particles. Therefore, platelets contain S100A8/A9 in membrane-enclosed vesicles and transfer it to target cells including endothelial cells and their precursors[53]. Probably, S100A8/A9-rich platelet-derived vesicles can play a pivotal role in developing endothelial dysfunction upon the manifestation of CV disease. Additionally, platelet-derived vesicles deliver oxidized lipids, P-selectin/P-selectin glycoprotein ligand 1, CD40 ligand (CD154) and induce B-cell synthesis and secretion of antigen-specific immunoglobulin G and potentiate the adaptive immune response through CD4+ cells[54-56]. Although restored endothelial cell junctions and vascular integrity are essential for prevention coagulation and thrombosis, the balance between promoting coagulation and mitigating endothelial dysfunction and vascular permeability has to turn to the side of potentiation of platelet activation status and pro-coagulant activity[57]. This effect appears to be harmful to majority patients with established CV diseases including asymptomatic atherosclerosis, acute coronary syndrome/myocardial infarction, hypertension, hypertensive crisis, heart failure, and peripheral artery disease.
Platelet-derived vesicles in hypertension
Platelet activation is common for arterial hypertension associating with increased P-selectin expression in the platelet membrane and elevated a circulating number of platelet-derived vesicles enriched phosphatidylserine that accompanies pro-coagulant blood activity[46]. It is been postulated that activation status of platelets in patients with uncontrolled arterial hypertension is predominantly a result of elevated shear force, low-grade systemic inflammation and increased sympathetic activity[47]. Yet, there is evidence that preeclampsia/eclampsia negatively impacts on the cerebral circulation and that this effect is associated with activated platelets and platelet-derived vesicles[48]. Moreover, placental and maternal platelet-derived EVs participate in critical signaling mechanism for feto-placental development and impairment in releasing and contents of vesicles in biological fluids depending on their origin is considered in the context of the initiation and progression of the pathogenesis of preeclampsia/eclampsia[49]. In fact, non-pregnant women with the risk factors of preeclampsia/eclampsia (overweight/obesity, diabetes mellitus, rheumatic disease, systemic vasculitis, dyslipidemia, anemia) as well as individuals with arterial hypertension were identified to have elevated levels of circulating platelet-derived vesicles[58-60]. It has been suggesting that circulating platelet-derived vesicles can induce vascular dysfunction, and through immune modulation they mediate thrombotic potency of plasma and remodeling of the vasculature. This mechanism appears to be common for arterial hypertension and preeclampsia, although initial triggers for both conditions can be distinguished (i.e., shear stress and sympathetic activity for hypertension and hypoxia/ischemia, damage of placenta villi for preeclampsia). Despite these differences, the final result of platelet activation and releasing of platelet-derived vesicles remain similar and it is considered an endothelial dysfunction, vascular remodeling, and pro-coagulation state, which affect target organ damage and contribute to poor clinical outcomes. However, renal denervation, which was able to sufficiently reduce sympathetic activity in patients with resistant hypertension, leads to a reduction of platelet activation, monocyte-platelet aggregates and a tendency to lowering circulating number of activated platelets in close relation to declined levels of several inflammatory markers, such as TNF-alpha, IL-1beta, IL-12, and monocyte chemoattractant protein-1[47,61]. Although there is strong interrelation between endothelial dysfunction in hypertension and spontaneous generation of platelet-derived vesicles, whether tight control for blood pressure will correspond to a significant decrease of platelet aggregation and reducing the number of circulating pro-coagulant platelet-derived vesicles is not fully understood[62]. However, diminishing of pro-coagulant activity of circulating platelet-derived vesicles can be achieved during effective antihypertensive therapy[62-64]. It has been noted that a wide range of antihypertensive drugs, such as calcium channel blockers, angiotensin-II receptor antagonists, angiotensin-converting enzyme inhibitors, are able to hamper the process of micro vesiculation and thereby to reduce a circulating number of platelet-derived vesicles[63,64]. Taken into consideration the role of the vesicles in the pathogenesis of hypertension a variable ability of several antihypertensive drugs to modify shaping and releasing of the particles onto circulation is showed to have promising and require more investigations in the future.
Platelet-derived vesicles in multifocal atherosclerosis
Previous studies have shown that platelet-derived vesicles directly mediated a formation of foam cells in atherosclerotic plaque shaping vulnerable plaque[65-67]. Therefore, increased levels of circulating TSP1+/CD142+ platelet-derived vesicles were found in older age individuals without CV disease, as well as in the younger individuals with high CV risk due to subclinical atherosclerosis[68,69] and patients with known CV diseases[70,71]. In fact, platelet-derived vesicles, which were enriched by phospholipids, correspond to atherosclerosis; vascular injury and plaque rupture acting as a trigger of microvascular inflammation, thrombosis, and endothelial dysfunction. However, the role of the particles in the clinical presentation of the disease requires to be clearly elucidated in large studies.
Platelet-derived vesicles in acute coronary syndrome and acute myocardial infarction
Acute coronary syndrome (ACS) and acute myocardial infarction (AMI) have been shown to be associated with an activation of various circulating cells, including platelets. Platelets are able to aggregate each other and with classical monocytes/macrophages shaping thrombus, embedding onto sub-cap space in a plaque, destroy a shoulder zone of the plaque and occlude the artery[72]. Being higher than in healthy individuals, aggregated platelets seem to consist of several cells (monocytes, macrophages, endothelial cells, and erythrocytes) and vesicles of various origin, such as phospholipid-rich platelet-derived extracellular vesicles expressed P-selectin (CD63), CD31, CD41a, and tissue factor (TF, CD142), elevated levels of which in peripheral blood strongly correspond to infarct size and in-hospital complications in AMI including mortality[73-75]. Moreover, ST-elevation AMI patients had higher levels of platelet-derived vesicles with immune phenotypes (CD61+/AV+, CD31+/AV+, CD42b+/AV+ and CD31+/CD42b+/AV+) compared to non-ST elevation AMI (STEMI) patients[76]. Yet, the investigator found that STEMI patients with NYHA class III heart failure (HF) had higher levels of circulating CD142+/AV+, CD14+/AV+ and CD14+/CD142+/AV+ endothelial cell-derived EVs as well as CD61+ platelet-derived vesicles than those in class I/II HF[76]. Thus, beyond biomarkers of cell activation, platelet-derived vesicles have numerous functional effects on the development of damaged vessel wall-induced arterial thrombi and blood thrombogenicity on areas of arterial damage contributing to atherothrombotic events. Indeed, phospholipids-rich platelets-derived vesicles have found the increased level of platelet-derived vesicles in survivors of AMI correlated well with thrombosis and soluble CD40 ligand (CD154)[77]. Authors concluded that the independent association between large platelet-derived vesicles and thrombin generation supports the concept that the formation of platelet-derived vesicles is important for increased coagulation activation in AMI patients[77]. Interestingly, the platelet-derived vesicles are discussed not the only factor directly mediating endothelial dysfunction and atherosclerosis, but they contribute to vascular reparation acting as factors of ischemic preconditioning on myocardial ischemia/reperfusion and vascular injury in correspondence with endothelial precursors[78]. Other studies reported that size of STEMI, lactate dehydrogenase activity, oxidative stress activity and the number of apoptotic cardiomyocytes were strongly and inversely associated with a number of platelet-derived vesicles in blood collected from the infarct-dependent artery[79,80]. Moreover, platelet-derived vesicles turn to be able to reduce the activity of caspase 3 and the expression of endoplasmic reticulum stress markers in the myocardium[79]. Additionally, endothelial precursors consolidate their interaction with platelets under dynamic flow conditions through secretion of platelet-derived vesicles and thereby mediate their regenerative potential[81-83]. Previously it has been reported that platelet-derived vesicles may modulate biological functions of hematopoietic cells and that they play an important but as yet not fully understood role in intercellular cross-talk in hematopoiesis and regeneration[84]. There is evidence that the platelet-derived vesicles can augment re-endothelialization capacity of circulating angiogenic cells[85]. All these facts support the hypothesis that angiopoietic capability of platelet-derived vesicles can modulate cardiac protection in ACS/STEMI[86,87]. Taken together, these findings open new insight on cardiac protection in ACS/STEMI based on an attenuation of endoplasmic reticulum stress apoptosis and angiogenesis through platelet-derived vesicles-endothelial precursors cooperation.
Platelet-derived vesicles in dyslipidemia and hypercholesterolemia
Dyslipidaemia is frequently associated with overweight, abdominal obesity, insulin resistant, metabolic syndrome, and type 2 diabetes mellitus. Increased levels of circulating platelet-derived vesicles were found in subjects with several metabolic diseases including metabolic syndrome and diabetes mellitus, in close relationship with severity of dyslipidemia and oxidative stress activity[88,89]. However, the role of hypercholesterolemia/dyslipidemia and hyperglycemia appears to be challenged. Hyperglycaemia is positively associated with the levels of circulating platelet-derived vesicles, whereas dyslipidemia influenced predominantly numbers of circulating endothelial cell-derived and erythrocyte-derived vesicles. Interestingly, the oxidative stress markers, such as plasma glutathione peroxidase and urinary 8-iso-prostaglandin F2 alpha, independently impacted on levels of platelet-derived vesicles[89]. Interestingly, the leptin can modulate platelet aggregation, shape platelet vesiculation and thereby links abdominal obesity and coagulopathy[90,91]. Moreover, leptin resistance rather than insulin resistance was found as a trigger for platelet activation that corresponds to increased risk of CV disease and CV events in patients with abdominal obesity, but not with diabetes mellitus[92,93]. There is evidence that the increased levels of low-density lipoprotein (LDL) cholesterol and oxidized LDL are able to induce membrane vesiculation and stimulate platelet-derived vesicle formation determining a coagulation potency of the plasma and altered endothelial-dependent vasodilation in patients with dyslipidemia[93-97]. Probably, phospholipid-rich platelet-derived vesicles influence directly and indirectly on vascular integrity and function in patients with hypercholesterolemia/dyslipidemia and their effects are mediated through several mechanisms including pro-coagulation state, impairment function of endothelial precursors, which play a pivotal role in vascular reparation, and accelerating atherosclerosis[93-96]. Thus, levels of a circulating number of platelet-derived vesicles may be a novel surrogate marker for the clinical evaluation of vascular damage in dyslipidemia state. Yet, the dynamic changes in platelet-derived profile could a promising biomarker for the identification of pleiotropic effects in lipid-lowering drugs.
Platelet-derived vesicles in atrial fibrillation
Atrial fibrillation (AF) is the most common cardiac arrhythmia, which is strongly associated with heart failure, sudden death, the pro-coagulant state and thromboembolic events including stroke, pulmonary thromboembolism[98]. Recent studies have shown that AF patients had significantly higher levels of total circulating vesicles originated from various cells including platelets to other individuals without AF and healthy volunteers[99-101]. It has found that numerous mechanisms that are associated with AF developing, such as hypoxia/ischemia, low or oscillatory shear stress, stretching, high ventricular heart rate, systemic low-grading inflammation and oxidative stress, are potent inducers of apoptotic cell death, which leads to the shedding of pro-coagulant vesicles within the vasculature[100]. Phosphatidylserine positive platelet-derived vesicles were found in elevated concentrations in peripheral blood collected from patients with a permanent form of AF while preserving sinus rhythm in post-AF/pre-AF periods in a persistent form of AF was associated with increased levels of platelet-derived vesicles with pro-coagulant capacity, but sufficiently lower in comparison with permanent AF patients[99,101,102]. However, the clinical studies have revealed that decreased apoptotic and lipid-rich platelet-derived vesicles after the restoration of sinus rhythm might enable restoration of hemostasis and reduce the risk of stroke and several thromboembolisms[101,102]. Conversely, there is evidence regarding that the platelet activation and its ability to release pro-coagulant P-selectin/phosphatidylcholine-rich vesicles in AF patients is a result of underlying CV diseases rather than the AF per se[102-104]. Interestingly, acetylsalicylic acid-clopidogrel combination therapy failed to reduce circulating biomarkers of thrombogenesis and platelet activation including a number of platelet-derived vesicles in patients with any form of AF. On the contrary, oral anticoagulants, such as vitamin K antagonists and novel oral anticoagulants, appear to be superior to combination acetylsalicylic acid-clopidogrel therapy in suppression platelet vesiculation and thereby in the prevention of AF-related thrombotic complication[105,106]. Additional study is required to further elucidate the core role of circulating platelet-derived vesicles in the pathophysiology of AF and its complications.
Platelet-derived vesicles in heart failure
HF are at higher risk of developing thrombosis and thromboembolic events[107]. It was found that progression of HF-related to developing “hypercoagulable phenotype”, which is associated with increased thrombin generation, decreased sensitivity to activated protein C, and over-production of pro-coagulant platelet-derived vesicles[108]. Moreover, in-hospital stay and mortality due to acute HF and acutely decompensated HF independently corresponded to a number of circulating platelet-derived vesicles[109]. However, there is limiting number of clinical studies that clearly elucidate the role of platelet-derived vesicles as a biological marker with predictive value, while other vesicles released from endothelial progenitors and mononuclears had exhibited prognostic utility in HF[110,111].
Platelet-derived vesicles in stroke
Thrombosis and platelet aggregation is defined as a crucial element in the pathophysiology of ischemic stroke and perpetrating brain ischemic event including transient ischemic attack (TIA)[112]. Despite platelet-derived vesicles, which had pro-coagulant potency, were found the most abundant in the circulation in patients with acute ischemic stroke and TIA, the predictive value for these findings is not still defined and required to be investigated in detail[113]. Obviously, perpetuating thrombosis in intracranial arteries could have a strong association with clinical outcomes and neurology deficiency, but structure, immune phenotypes, and number of platelet-derived vesicles were accompanied to vascular tone, levels of inflammatory cytokine, thrombus formation and oxidative stress biomarkers, while pro-coagulant platelet-derived vesicles may substantially contribute to ischemic brain disease in several settings, such as neuroinflammatory conditions[114,115]. Whether platelet-derived vesicle profile corresponds to the severity of intracranial hemorrhage, subarachnoid hemorrhage, and intracranial vasospasm is not fully understood. Probably, this assumption should address the future.
Conclusion
Although the innate mechanisms regarding the regulation of platelet-derived vesicle secretion and impact on target cells are not fully elucidated and required more investigations, increased content of platelet-derived vesicles, even in normal blood conditions, may enhance vascular damage, microvascular obstruction, and ongoing thrombus formation. It is suggested that platelet-derived vesicles may be considered a potential biological marker for endothelial dysfunction, thrombotic/thromboembolic events and progression of atheroma and may be implicated in the pathogenesis of coagulation abnormalities encountered in patients with known CV diseases and CV events. Additionally, a number of circulating platelet-derived vesicles may be represented as a useful CV risk stratification tool as well as a novel therapeutic target for anti-thrombotic drugs.
Declarations
Authors’ contributionsConception and design of the study: Berezin AE, Berezin AA
Performed data searching and interpretation: Berezin AE, Berezin AA
Manuscript writing: Berezin AE, Berezin AA
Provided administrative, technical, and material support: Berezin AA
Availability of data and materialsNot applicable.
Financial support and sponsorshipNone.
Conflicts of interestAll authors declare that there are no conflicts of interest.
Ethical approval and consent to participateNot applicable.
Consent for publicationNot applicable.
Copyright© The Author(s) 2019.
REFERENCES
1. Younus A, Aneni EC, Spatz ES, Osondu CU, Roberson L, et al. A systematic review of the prevalence and outcomes of ideal cardiovascular health in US and non-US populations. Mayo Clin Proc 2016;91:649-70.
2. Sotos-Prieto M, Mattei J, Cook NR, Hu FB, Willett WC, et al. Association between a 20-year cardiovascular disease risk score based on modifiable lifestyles and total and cause-specific mortality among US men and women. J Am Heart Assoc 2018;7:e010052.
3. Peters SAE, Muntner P, Woodward M. Sex differences in the prevalence of, and trends in, cardiovascular risk factors, treatment, and control in the United States, 2001 to 2016. Circulation 2019;139:1025-35.
4. Bress AP, Colantonio LD, Cooper RS, Kramer H, Booth JN 3rd, et al. Potential cardiovascular disease events prevented with adoption of the 2017 American college of cardiology/American heart association blood pressure guideline. Circulation 2019;139:24-36.
5. Leong DP, Joseph PG, McKee M, Anand SS, Teo KK, et al. Reducing the global burden of cardiovascular disease, part 2: prevention and treatment of cardiovascular disease. Circ Res 2017;121:695-710.
6. Aggarwal M, Aggarwal B, Rao J. Integrative medicine for cardiovascular disease and prevention. Med Clin North Am 2017;101:895-923.
7. Shanmuganathan M, Vughs J, Noseda M, Emanuel C. Exosomes: basic biology and technological advancements suggesting their potential as ischemic heart disease therapeutics. Front Physiol 2018;9:1159.
8. Berezin A, Zulli A, Kerrigan S, Petrovic D, Kruzliak P. Predictive role of circulating endothelial-derived microparticles in cardiovascular diseases. Clin Biochem 2015;48:562-8.
9. Nawaz M, Shah N, Zanetti BR, Maugeri M, Silvestre RN, et al. Extracellular vesicles and matrix remodeling enzymes: the emerging roles in extracellular matrix remodeling, progression of diseases and tissue repair. Cells 2018;7:E167.
10. La Marca V, Fierabracci A. Insights into the Diagnostic potential of extracellular vesicles and their miRNA signature from liquid biopsy as early biomarkers of diabetic micro/macrovascular complications. Int J Mol Sci 2017;18:E1974.
11. Berezin AE, Kremzer AA, Berezina TA, Martovitskaya YV. The pattern of circulating microparticles in chronic heart failure patients with metabolic syndrome: Relevance to neurohumoral and inflammatory activation. BBA Clin 2015;4:69-75.
12. Caporali A, Martello A, Miscianinov V, Maselli D, Vono R, et al. Contribution of pericyte paracrine regulation of the endothelium to angiogenesis. Pharmacol Ther 2017;171:56-64.
13. De Toro J, Herschlik L, Waldner C, Mongini C. Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front Immunol 2015;6:203.
15. Berezin AE, Kremzer AA, Cammarota G, Zulli A, Petrovic D, et al. Circulating endothelial-derived apoptotic microparticles and insulin resistance in non-diabetic patients with chronic heart failure. Clin Chem Lab Med 2016;54:1259-67.
16. Chen BY, Sung CW, Chen C, Cheng CM, Lin DP, et al. Advances in exosomes technology. Clin Chim Acta 2019;493:14-9.
17. Juan T, Fürthauer M. Biogenesis and function of ESCRT-dependent extracellular vesicles. Semin Cell Dev Biol 2018;74:66-77.
18. Hervera A, Santos CX, De Virgiliis F, Shah AM, Di Giovanni S. Paracrine mechanisms of redox signalling for postmitotic cell and tissue regeneration. Trends Cell Biol 2019; doi: 10.1016/j.tcb.2019.01.006.
19. Mobarak H, Heidarpour M, Lolicato F, Nouri M, Rahbarghazi R, et al. Physiological impact of extracellular vesicles on the female reproductive system; highlights possible restorative effects on female age-related fertility. Biofactors 2019; doi: 10.1002/biof.1497.
20. Navarro A, Molins L, Marrades RM, Moises J, Viñolas N, et al. Exosome Analysis in Tumor-Draining Pulmonary Vein Identifies NSCLC Patients with Higher Risk of Relapse after Curative Surgery. Cancers (Basel) 2019;11:E249.
21. Aslan C, Maralbashi S, Salari F, Kahroba H, Sigaroodi F, et al. Tumor-derived exosomes: Implication in angiogenesis and antiangiogenesis cancer therapy. J Cell Physiol 2019; doi: 10.1002/jcp.28374.
22. Yang L, Zhu J, Zhang C, Wang J, Yue F, et al. Stem cell-derived extracellular vesicles for myocardial infarction: a meta-analysis of controlled animal studies. Aging (Albany NY) 2019;11:1129-50.
23. Bei Y, Das S, Rodosthenous RS, Holvoet P, Vanhaverbeke M, et al. Extracellular vesicles in cardiovascular theranostics. Theranostics 2017;7:4168-82.
24. Connor DE, Exner T, Ma DD, Joseph JE. The majority of circulating platelet-derived microparticles fail to bind annexin V, lack phospholipid-dependent pro-coagulant activity and demonstrate greater expression of glycoprotein Ib. Thromb Haemost 2010;103:1044-52.
25. Biró E, Akkerman JW, Hoek FJ, Gorter G, Pronk LM, Sturk A, et al. The phospholipid composition and cholesterol content of platelet-derived microparticles: a comparison with platelet membrane fractions. J Thromb Haemost 2005;3:2754-63.
26. Diehl P, Fricke A, Sander L, Stamm J, Bassler N, et al. Microparticles: major transport vehicles for distinct microRNAs in circulation. Cardiovasc Res 2012;93:633-44.
27. Habersberger J, Strang F, Scheichl A, Htun N, Bassler N, et al. Circulating microparticles generate and transport monomeric C-reactive protein in patients with myocardial infarction. Cardiovasc Res 2012;96:64-72.
28. Forlow SB, McEver RP, Nollert MU. Leukocyte-leukocyte interactions mediated by platelet microparticles under flow. Blood 2000;95:1317-23.
29. Dinkla S, van Cranenbroek B, van der Heijden WA, He X, Wallbrecher R, et al. Platelet microparticles inhibit IL-17 production by regulatory T cells through P-selectin. Blood 2016;127:1976-86.
30. Baker N, O’Meara SJ, Scannell M, Maderna P, Godson C. Lipoxin A4: anti-inflammatory and anti-angiogenic impact on endothelial cells. J Immunol 2009;182:3819-26.
31. Kim HK, Song KS, Chung JH, Lee KR, Lee SN. Platelet microparticles induce angiogenesis in vitro. Br J Haematol 2004;124:376-84.
32. Kenari AN, Kastaniegaard K, Greening DW, Shambrook M, Stensballe A, et al. Exosome-mimetic nanovesicles contain distinct proteome and post-translational modified protein cargo, in comparison to exosomes. Proteomics 2019;19:e1800161.
33. Vagner T, Chin A, Mariscal J, Bannykh S, Engman D, et al. Protein composition reflects extracellular vesicle heterogeneity. Proteomics 2019;19:e1800167.
34. van der Pol E, Böing AN, Gool EL, Nieuwland R. Recent developments in the nomenclature, presence, isolation, detection and clinical impact of extracellular vesicles. J Thromb Haemost 2016;14:48-56.
35. Zaldivia MTK, McFadyen JD, Lim B, Wang X, Peter K. Platelet-derived microvesicles in cardiovascular diseases. Front Cardiovasc Med 2017;4:74.
36. Aatonen M, Valkonen S, Böing A, Yuana Y, Nieuwland R, et al. Isolation of platelet-derived extracellular vesicles. Methods Mol Biol 2017;1545:177-88.
37. Hedley BD, Llewellyn-Smith N, Lang S, Hsia CC, MacNamara N, et al. Combined accurate platelet enumeration and reticulated platelet determination by flow cytometry. Cytometry B Clin Cytom 2015;88:330-7.
39. Clemetson KJ, Clemetson JM. Platelet GPIb complex as a target for anti-thrombotic drug development. Thromb Haemost 2008;99:473-9.
40. Chevillet JR, Kang Q, Ruf IK, Briggs HA, Vojtech LN, Hughes SM, et al. Quantitative and stoichiometric analysis of the microRNA content of exosomes. Proc Natl Acad Sci U S A 2014;111:14888-93.
41. van der Pol E, Boing AN, Harrison P, Sturk A, Nieuwland R. Classification, functions, and clinical relevance of extracellular vesicles. Pharmacological Reviews 2012;64:676-705.
42. Liu C, Zhao J, Tian F, Chang J, Zhang W, Sun J. λ-DNA and aptamer mediated sorting and analysis of extracellular vesicles. J Am Chem Soc 2019;141:3817-21.
44. Lacroix R, Robert S, Poncelet P, Kasthuri R, Key N, et al. Standardization of platelet-derived microparticle enumeration by flow cytometry with calibrated beads: results of the international society on thrombosis and haemostasis SSC collaborative workshop. J Thromb Haemost 2010;8:2571-4.
45. Tao SC, Guo SC, Zhang CQ. Platelet-derived extracellular vesicles: an emerging therapeutic approach. Int J Biol Sci 2017;13:828-34.
46. Zaldivia MTK, Hering D, Marusic P, Sata Y, Lee R, et al. Successful renal denervation decreases the platelet activation status in hypertensive patients. Cardiovasc Res 2019; doi: 10.1093/cvr/cvz033.
47. Zaldivia MT, Rivera J, Hering D, Marusic P, Sata Y, et al. Renal denervation reduces monocyte activation and monocyte-platelet aggregate formation: an anti-inflammatory effect relevant for cardiovascular risk. Hypertension 2017;69:323-31.
48. Barnes JN, Harvey RE, Miller KB, Jayachandran M, Malterer KR, et al. Cerebrovascular reactivity and vascular activation in postmenopausal women with histories of preeclampsia. Hypertension 2018;71:110-7.
49. Van Wijk MJ, Van Bavel E, Sturk A, Nieuwland R. Microparticles in cardiovascular diseases. Cardiovasc Res 2003;59:277-87.
50. Yano Y, Kambayashi J, Shiba E, Sakon M, Oiki E, et al. The role of protein phosphorylation and cytoskeletal reorganization in microparticle formation from the platelet plasma membrane. Biochem J 1994;299:303-8.
51. Gemmel CH, Sefton MV, Yeo E. Platelet-derived microparticle formation involves glycoprotein IIb-IIIa. Inhibition by RGDS and a Glanzmann’s thrombasthenia defect. J Biol Chem 1993;268:14586-9.
52. Cauwenberghs S, Feijge MA, Harper AG, Sage SO, Curvers J, Heemskerk JW. Shedding of procoagulant microparticles from unstimulated platelets by integrin-mediated destabilization of the actin cytoskeleton. FEBS Lett 2006;580:5313-20.
53. Nomura S, Komiyama Y, Miyake T, Miyazaki Y, Kido H, et al. Amyloid-protein precursor-rich platelet microparticles in thrombotic disease. Thromb Haemost 1994;72:519-22.
54. Furie B, Furie BC. Role of platelet P-selectin and microparticle PSGL-1 in thrombus formation. Trends Mol Med 2004;10:171-8.
55. Barry OP, Practico D, Lawson JA, FitzGerald GA. Transcellular activation of platelets and endothelial cells by bioactive lipids in platelet microparticles. J Clin Invest 1997;99:2118-27.
56. Garcia B, Smalley DM, Cho H, Shabanowitz J, Ley K, Hunt DF. The platelet particle proteome. J Proteome Res 2005;4:1516-21.
57. Sprague DL, Elzey BD, Crist SA, Waldschmidt TJ, Jensen RJ, et al. Platelet-mediated modulation of adaptive immunity: unique delivery of CD154 signal by platelet-derived membrane vesicles. Blood 2008;111:5028-36.
58. Gilani SI, Weissgerber TL, Garovic VD, Jayachandran M. Preeclampsia and extracellular vesicles. Curr Hypertens Rep 2016;18:68.
59. Stepanian A, Bourguignat L, Hennou S, Coupaye M, Hajage D, Salomon L, et al. Microparticle increase in severe obesity: not related to metabolic syndrome and unchanged after massive weight loss. Obesity (Silver Spring) 2013;21:2236-43.
60. Preston RA, Jy W, Jimenez JJ, Mauro LM, Horstman LL, Valle M, et al. Effects of severe hypertension on endothelial and platelet microparticles. Hypertension 2003;41:211-7.
61. Sabatier F, Darmon P, Hugel B, Combes V, Sanmarco M, Velut JG, et al. Type 1 and type 2 diabetic patients display different patterns of cellular microparticles. Diabetes 2002;51:2840-5.
62. Bacha NC, Levy M, Guerin CL, Le Bonniec B, Harroche A, et al. Treprostinil treatment decreases circulating platelet microvesicles and their procoagulant activity in pediatric pulmonary hypertension. Pediatr Pulmonol 2019;54:66-72.
63. Rosińska J, Łukasik M, Kozubski W. The impact of vascular disease treatment on platelet-derived microvesicles. Cardiovasc Drugs Ther 2017;31:627-44.
64. Berkels R, Egink G, Marsen TA, Bartels H, Roesen R, et al. Nifedipine increases endothelial nitric oxide bioavailability by antioxidative mechanisms. Hypertension 2001;37:240-5.
65. Labiós M, Martínez M, Gabriel F, Guiral V, Ruiz Aja S, et al. Effect of eprosartan on cytoplasmic free calcium mobilization, platelet activation, and microparticle formation in hypertension. Am J Hypertens 2004;17:757-63.
66. Miyazawa B, Trivedi A, Togarrati PP, Potter D, Baimukanoya G, et al. Regulation of endothelial cell permeability by platelet-derived extracellular vesicles. J Trauma Acute Care Surg 2019; doi: 10.1097/TA.0000000000002230.
67. Nomura S, Suzuki M, Katsura K, et al. Platelet-derived microparticles may influence the development of atherosclerosis in diabetes mellitus. Atherosclerosis 1995;116:235-40.
68. Akbiyik F, Ray DM, Gettings KF, Blumberg N, Francis CW, Phipps RP. Human bone marrow megakaryocytes and platelets express PPARgamma, and PPARgamma agonists blunt platelet release of CD40 ligand and thromboxanes. Blood 2004;104:1361-8.
69. Burnouf T, Goubran HA, Chou ML, Devos D, Radosevic M. Platelet microparticles: detection and assessment of their paradoxical functional roles in disease and regenerative medicine. Blood Rev 2014;28:155-66.
70. Tan KT, Lip GY. The potential role of platelet microparticles in atherosclerosis. Thromb Haemost 2005;94:488-92.
71. Suades R, Padró T, Alonso R, Mata P, Badimon L. High levels of TSP1+/CD142+ platelet-derived microparticles characterise young patients with high cardiovascular risk and subclinical atherosclerosis. Thromb Haemost 2015;114:1310-21.
72. van der Zee PM, Biró E, Ko Y, de Winter RJ, Hack CE, et al. P-selectin- and CD63-exposing platelet microparticles reflect platelet activation in peripheral arterial disease and myocardial infarction. Clin Chem 2006;52:657-64.
73. Loguinova M, Pinegina N, Kogan V, Vagida M, Arakelyan A, et al. Monocytes of Different Subsets in Complexes with Platelets in Patients with Myocardial Infarction. Thromb Haemost 2018;118:1969-81.
74. Min PK, Kim JY, Chung KH, Lee BK, Cho M, et al. Local increase in microparticles from the aspirate of culprit coronary arteries in patients with ST-segment elevation myocardial infarction. Atherosclerosis 2013;227:323-8.
75. Vagida MS, Arakelyan A, Lebedeva AM, Grivel JC, Shpektor AV, et al. Analysis of Extracellular Vesicles Using Magnetic Nanoparticles in Blood of Patients with Acute Coronary Syndrome. Biochemistry (Mosc) 2016;81:382-91.
76. Jung C, Sörensson P, Saleh N, Arheden H, Rydén L, et al. Circulating endothelial and platelet derived microparticles reflect the size of myocardium at risk in patients with ST-elevation myocardial infarction. Atherosclerosis 2012;221:226-31.
77. Chiva-Blanch G, Laake K, Myhre P, Bratseth V, Arnesen H, et al. Platelet-, monocyte-derived and tissue factor-carrying circulating microparticles are related to acute myocardial infarction severity. PLoS One 2017;12:e0172558.
78. Suades R, Padró T, Vilahur G, Badimon L. Circulating and platelet-derived microparticles in human blood enhance thrombosis on atherosclerotic plaques. Thromb Haemost 2012;108:1208-19.
79. Michelsen AE, Brodin E, Brosstad F, Hansen JB. Increased level of platelet microparticles in survivors of myocardial infarction. Scand J Clin Lab Invest 2008;68:386-9.
80. Liu M, Wang Y, Zhu Q, Zhao J, Wang Y, et al. Protective effects of circulating microvesicles derived from ischemic preconditioning on myocardial ischemia/reperfusion injury in rats by inhibiting endoplasmic reticulum stress. Apoptosis 2018;23:436-48.
81. Porto I, Biasucci LM, De Maria GL, Leone AM, Niccoli G, et al. Intracoronary microparticles and microvascular obstruction in patients with ST elevation myocardial infarction undergoing primary percutaneous intervention. Eur Heart J 2012;33:2928-38.
82. Alexandru N, Andrei E, Dragan E, Georgescu A. Interaction of platelets with endothelial progenitor cells in experimental atherosclerosis: Role of transplanted endothelial progenitor cells and platelet microparticles. Biol Cell 2015;107:189-204.
83. Berezin AE. Are endothelial cell-derived microparticles predictive biomarkers in cardiovascular diseases? Atherosclerosis: open Access 2015;1:e101-3.
84. Mause SF, Ritzel E, Liehn EA, Hristov M, Bidzhekov K, et al. Platelet microparticles enhance the vasoregenerative potential of angiogenic early outgrowth cells after vascular injury. Circulation 2010;122:495-506.
85. Baj-Krzyworzeka M, Majka M, Pratico D, Ratajczak J, Vilaire G, et al. Platelet-derived microparticles stimulate proliferation, survival, adhesion, and chemotaxis of hematopoietic cells. Exp Hematol 2002;30:450-9.
86. Ohtsuka M, Sasaki K, Ueno T, Seki R, Nakayoshi T, et al. Platelet-derived microparticles augment the adhesion and neovascularization capacities of circulating angiogenic cells obtained from atherosclerotic patients. Atherosclerosis 2013;227:275-82.
87. Chen BA, Zhong YJ, Huang CY, Li CP, Shi GY, et al. Effects of platelet-derived membrane microparticles on angiogenesis in chick chorioallantoic membranes. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2007;15:1070-3.
88. Ma F, Liu H, Shen Y, Zhang Y, Pan S. Platelet-derived microvesicles are involved in cardioprotective effects of remote preconditioning. Int J Clin Exp Pathol 2015;8:10832-9.
89. Helal O, Defoort C, Robert S, Marin C, Lesavre N, et al. Increased levels of microparticles originating from endothelial cells, platelets and erythrocytes in subjects with metabolic syndrome: relationship with oxidative stress. Nutr Metab Cardiovasc Dis 2011;21:665-71.
90. Connolly KD, Willis GR, Datta Dev BN, et al. Lipoprotein apheresis reduces circulating microparticles in individuals with familial hypercholesterolemia. J Lipid Res 2014;55:2064-72.
91. Konstantinides S, Schafer K, Koschnick S, Loskutoff DJ. Leptin-dependent platelet aggregation and arterial thrombosis suggest a mechanism for atherothrombotic disease in obesity. J Clin Invest 2001;108:1533-40.
92. Nakata M, Yada T, Soejima N, Maruyama I. Leptin promotes aggregation of human platelets via the long form of its receptor. Diabetes 1999;48:426-9.
93. Lukasik M, Michalak S, Dworacki G, Kaczmarek M, Watala C, et al. Reactive leptin resistance and the profile of platelet activation in acute ischaemic stroke patients. Thromb Haemost 2012;108:107-18.
94. Nomura S, Shouzu A, Omoto S, Nishikawa M, Iwasaka T, et al. Activated platelet and oxidized LDL induce endothelial membrane vesiculation: clinical significance of endothelial cell-derived microparticles in patients with type 2 diabetes. Clin Appl Thromb Hemost 2004;10:205-15.
95. Tushuizen ME, Nieuwland R, Scheffer PG, Sturk A, Heine RJ, Diamant M. Two consecutive high-fat meals affect endothelial-dependent vasodilation, oxidative stress and cellular microparticles in healthy men. J Thromb Haemost 2006;4:1003-10.
96. Heinrich LF, Andersen DK, Cleasby ME, Lawson C. Long-term high-fat feeding of rats results in increased numbers of circulating microvesicles with pro-inflammatory effects on endothelial cells. Br J Nutr 2015;113:1704-11.
97. Pirro M, Schillaci G, Paltriccia R, Bagaglia F, Menecali C, et al. the Increased ratio of CD31+/CD42- microparticles to endothelial progenitors as a novel marker of atherosclerosis in hypercholesterolemia. Arterioscler Thromb Vasc Biol 2006;26:2530-5.
98. Conway DS, Pearce LA, Chin BS, Hart RG, Lip GY. Plasma von Willebrand factor and soluble p-selectin as indices of endothelial damage and platelet activation in 1321 patients with nonvalvular atrial fibrillation: relationship to stroke risk factors. Circulation 2002;106:1962-7.
99. Siwaponanan P, Keawvichit R, Udompunturak S, Hunnangkul S, Reesukumal K, et al. Altered profile of circulating microparticles in nonvalvular atrial fibrillation. Clin Cardiol 2019;42:425-31.
100. Jesel L, Abbas M, Toti F, Cohen A, Arentz T, et al. Microparticles in atrial fibrillation: a link between cell activation or apoptosis, tissue remodeling and thrombogenicity. Int J Cardiol 2013;168:660-9.
101. Horstman LL, Jy W, Bidot CJ, Nordberg ML, Minagar A, et al. Potential roles of cell-derived microparticles in ischemic brain disease. Neurol Res 2009;31:799-806.
102. Choudhury A, Chung I, Blann AD, Lip GYH. Elevated platelet microparticle levels in nonvalvular atrial fibrillation: relationship to p-selectin and antithrombotic therapy. Chest 2007;131:809-15.
103. Choudhury A, Chung I, Blann AD, Lip GY. Platelet surface CD62P and CD63, mean platelet volume, and soluble/platelet P-selectin as indexes of platelet function in atrial fibrillation: a comparison of “healthy control subjects” and “disease control subjects” in sinus rhythm. J Am Coll Cardiol 2007;49:1957-64.
104. Tan KT, Tayebjee MH, Lim HS, Lip GY. Clinically apparent atherosclerotic disease in diabetes is associated with an increase in platelet microparticle levels. Diabet Med 2005;22:1657-62.
105. Kamath S, Blann AD, Chin BS, Lanza F, Aleil B, et al. A study of platelet activation in atrial fibrillation and the effects of antithrombotic therapy. Eur Heart J 2002;23:1788-95.
106. Choudhury A, Chung I, Blann A, Lip GY. Platelet adhesion in atrial fibrillation. Thromb Res 2007;120:623-9.
108. Popovic B, Zannad F, Louis H, Clerc-Urmès I, Lakomy C, et al. Endothelial-driven increase in plasma thrombin generation characterizing a new hypercoagulable phenotype in acute heart failure. Int J Cardiol 2019;274:195-201.
109. Roura S, Gálvez-Montón C, de Gonzalo-Calvo D, Valero AG, Gastelurrutia P, et al. Extracellular vesicles do not contribute to higher circulating levels of soluble LRP1 in idiopathic dilated cardiomyopathy. J Cell Mol Med 2017;21:3000-9.
110. Berezin AE. Prognostication in different heart failure phenotypes: the role of circulating biomarkers. J Circ Biomark 2016;5:6.
111. Berezin AE, Kremzer AA, Martovitskaya YV, Samura TA, Berezina TA. The predictive role of circulating microparticles in patients with chronic heart failure. BBA Clin 2014;3:18-24.
112. Arjmand S, Pardakhty A, Forootanfar H, Khazaeli P. A road to bring Brij52 back to attention: Shear stress sensitive Brij52 liposomal carriers for targeted drug delivery to obstructed blood vessels. Med Hypotheses 2018;121:137-41.
113. Viera AJ, Mooberry M, Key NS. Microparticles in cardiovascular disease pathophysiology and outcomes. J Am Soc Hypertens 2012;6:243-52.
114. Horstman LL, Jy W, Bidot CJ, Nordberg ML, Minagar A, et al. Potential roles of cell-derived microparticles in ischemic brain disease. Neurol Res 2009;31:799-806.
Cite This Article
Export citation file: BibTeX | RIS
OAE Style
Berezin AE, Berezin AA. Platelet-derived vesicles: diagnostic and predictive value in cardiovascular diseases. J Unexplored Med Data 2019;4:4. http://dx.doi.org/10.20517/2572-8180.2019.05
AMA Style
Berezin AE, Berezin AA. Platelet-derived vesicles: diagnostic and predictive value in cardiovascular diseases. Journal of Unexplored Medical Data. 2019; 4: 4. http://dx.doi.org/10.20517/2572-8180.2019.05
Chicago/Turabian Style
Berezin, Alexander E., Alexander A. Berezin. 2019. "Platelet-derived vesicles: diagnostic and predictive value in cardiovascular diseases" Journal of Unexplored Medical Data. 4: 4. http://dx.doi.org/10.20517/2572-8180.2019.05
ACS Style
Berezin, AE.; Berezin AA. Platelet-derived vesicles: diagnostic and predictive value in cardiovascular diseases. J. Unexplored. Med. Data. 2019, 4, 4. http://dx.doi.org/10.20517/2572-8180.2019.05
About This Article
Special Issue
Copyright

Data & Comments
Data


 Cite This Article 14 clicks
Cite This Article 14 clicks

 Like This Article 1
likes
Like This Article 1
likes






Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.