
Molecular characterization of cancer-associated fibroblasts isolated from human colorectal cancer as a major stromal cell component promoting metastasis

Abstract
Aim: Colorectal cancer (CRC) remains a disease with poor prognosis and limited overall 5-year survival rate, which highlights a clear clinical need for novel treatment strategies. The stromal cancer-associated fibroblasts (CAFs) strongly dictate the metastatic potential of CRC and hence warrant investigation for clinical outcome.
Methods: The authors established primary cultures of CAFs from metastatic CRC patients and performed cellular characterization using phase contrast microscopy and histological staining followed by light microscopy. The isolated CAFs were further explored for the molecular characterization of genes which promote the process of metastasis.
Results: The stromal CAFs maintain their fibroblastic phenotype in vitro and manifested high proliferation potential. To explore the gene expression profile, total RNA was isolated from the primary culture of CAFs and qualitative RT-PCR was performed. The cultured CAFs exhibited gene signatures associated with cancer stemness, epithelial to mesenchymal transition induction and inflammatory cytokines/chemokines favoring metastasis. These genes play a pivotal role in chemotherapy resistance and are also associated with poor prognosis in CRC.
Conclusion: This study further delineates the role of CAFs as a part of the “corrupted” stromal cells within the tumor microenvironment in establishing and orchestrating the metastatic fate of CRC. The gene expression profile clearly indicates that CAFs represent a potential target for improved therapy of CRC.
Keywords
Introduction
Colorectal cancer (CRC) is one of the most common cancers and a high dietary intake of red meat along with low-fiber food increases the risk.[1] The primary tumor often remains asymptomatic and subsequently results in metastasis, which is the leading cause of mortality in many patients.[2] Despite the recent advances in chemotherapy regimens, chemoresistance remains an obstacle and often leads to severe side-effects. Relapses occur in 15-20% of colorectal patients treated with chemotherapy.[3] Thus, the identification of prognostic markers should improve both the clinical outcome and effectiveness of treatment in such patients.
One of the most consistent features of CRC is the attainment of epithelial to mesenchymal transition (EMT), which is associated with poor prognosis and resistance to chemotherapy.[4] This phenotypic switching of epithelial-origin cancer cells to acquire mesenchymal traits typically involves acquisition of a cancer stem-cell state. However, the carcinoma genome is exposed to transcriptional signals originating not only from the cancer cells but also from the tumor microenvironment.[5] Albeit much less studied, the tumor microenvironment is composed of reactive stromal cells such as cancer associated fibroblasts (CAFs) which are crucial for colonizing tumors at secondary sites during metastasis. Earlier works have suggested that the pattern of gene expression in stromal cells constitutes a signature that predicts the metastatic potential of CRC and possibly clinical relevance.[6] Thus, it seems plausible that the gene expression trait in stromal cells dictates the malignant fate of CRC and hence warrants further investigation.
To gain further insight into the gene expression characteristics in stromal CAFs of CRC, we have proposed to isolate CAFs from patients with metastatic CRC to study the gene expression profile of these cells by RT-PCR. This study will report the stem-cell like characteristic of CAFs with concomitant expression of metastatic traits and oncogenes and thus reflects the clinical relevance of stromal gene expression changes in CRC. Since the stromal CAFs also dictate the malignant fate of CRC, they serve as one of the therapeutic targets to potentially block the cross-talk between cancer cells and stromal environment in order to halt disease progression and thereby possibly cure CRC.
Methods
Establishment of CAFs cells from peripheral blood of metastatic CRC patients
A metastatic colon cancer patient at stage IV attending the Oncology Department of Jaslok Hospital and Research Centre, Mumbai was recruited for this study as per the ethical guidelines of this hospital. Our laboratory is approved lab for molecular diagnostic cancer testing. Ten mL of peripheral blood was collected in an EDTA vacutainer and was layered on Histopaque (Sigma-Aldrich, St. Louis, MO) Ficoll gradient to isolate PBMCs. These isolated PBMCs were initially cultured in complete RPMI-1640 (Life Technologies, Inc., Grand Island, NY) growth medium containing 10% FBS (Fetal Bovine Serum, GIBCO), 1% penicillin-streptomycin (Life Technologies, Inc.) and 2 mmol/L L-glutamine (Life Technologies, Inc.) for 24 h. Following overnight incubation, all the unattached cells were removed and only the adhered cells were fed with complete DMEM medium containing 10% FBS, 1% penicillin-streptomycin and 2 mmol/L L-glutamine for almost 40-45 days by feeding with fresh growth media on alternate days. During 45 days of incubation, we found that most of other cells vanished from culture dishes. But after 40-45 days, an outgrowth of fibroblast-like cells appeared in these cultures and started growing very rapidly. These cells reached confluency within one week and were monitored regularly for their morphology using phase-contrast microscopy. After achieving confluency, these cells were passaged by using 0.25% PBS -trypsin and transferred to a larger flask for expansion of this cell line. These cells were then stored at -85 °C until further experimentation.
Histological staining and light microscopy of CAFs
The harvested CAFs in the earliest passages were seeded in 65-mm petri dishes (Thermo Scientific, Nunc Ltd. UK), incubated for 24 h, rinsed with PBS (Life Technologies, Inc., Grand Island, NY) and subjected to three different stains namely Giemsa stain (Sigma-Aldrich, St. Louis, MO), Alzarin Red (Merck Millipore, Darmstadt, Germany), Oil Red (Merck Millipore, Darmstadt, Germany) as described previously to study their morphological characteristics.[7] Cells were photographed under a light microscope equipped with TSview software (Tucsen Imaging, Fuzhou, China).
Cell growth kinetics of CAFs
A total of 4 × 104 cells of CAFs were seeded per well in a 6-well plate (Thermo Scientific, Nunc Ltd. UK) in 1 mL of DMEM medium containing 10% FBS, 1% penicillin-streptomycin and 2 mmol/L L-glutamine. The cells were incubated in a CO2 incubator at 37 °C with 5% CO2 for 1, 2, 3, 4, 5, and 6 days intervals. Cell growth was terminated at day 1, 2, 3, 4, 5, and 6 by washing cells with 1 × PBS and then trypsinising using 0.5 mL of 0.25% PBS-trypsin for 3 min following by addition of 0.5 mL growth medium to stop this reaction. Then cells were properly mixed and cells counted on a hemocytometer chamber using erythrosine B (Sigma-Aldrich, St. Louis, MO). The cell count was then expressed as count/mL of cells by the usual counting formulae, i.e. number of cells in 4 squares of hemocytometer divided by 4, and then multiplied by dilution factor and counting factor 104, giving the total count/mL of cells. The experiment was repeated 3 times allowing calculation of mean and standard deviation. A graph was plotted of cells per milliliter at each time point, namely 1, 2, 3, 4, 5, and 6 days. The intersection of two points in 24 h on this graph provides the cell growth rate of these cells.
RNA extraction and cDNA synthesis
For the extraction of total RNA from CAFs, the cultures in earliest passages were maintained for 48 h and treated with 500 µL of Trizol reagent (Invitrogen, Carlsbad, CA). RNA was harvested according to the manufacturer’s instructions and further quantified using spectrophotometer. Nearly 2 µg of total RNA was reverse transcribed to cDNA using a high capacity cDNA Reverse transcriptase kit (Applied Biosystem, CA).
RT-PCR for gene expression study
To gain further insight into the gene expression characteristics of CAFs, we investigated the expression of a fibroblast marker (vimentin); epithelial markers (keratin 18 and e-cadherin); pluripotency stemness markers (Oct4, Nanog, Sox2, Klf4); metastatic markers (MMP1, MMP2, MMP9, VEGF); chemokines (SDF-1, CXCR4, CXCR7); and cytokines (IL6, TNF-α). The RT-PCR condition was carried out using the following cycling conditions: the initial cDNA denaturation was carried out at 95 °C for 5 min, followed by 40 cycles of 95 °C for 30 s, and annealing temperatures were diversely set at 55 °C (Oct4, Nanog, keratin 18, MMP1), 57 °C (Sox2), 58 °C (MMP2, IL6, TNF-α), 60 °C (e-cadherin, SDF-1, CXCR4, CXCR7), 62 °C (vimentin, VEGF), 64 °C (MMP9, Klf4) for 30 s, and extension at 72 °C for 1 min. β actin was used as an internal housekeeping gene. Most of the sequences for the primers have already been described previously by Potdar’s group.[7,8]
Results
Primary culture of CAFs isolated from colorectal tumor displays features of activated fibroblasts
To characterize stromal CAFs in the microenvironment of CRC, fresh blood from the patient suffering from metastatic CRC was used for isolating primary CAFs. After 6-7 weeks of incubation, CAFs were seen outgrowing and remained attached to the culture plate [Figure 1]. All the unattached cells were removed through washing. The confluent monolayer of CAFs was subjected to trypsinization and expanded by a large amount for further experimentation. Cells were cryopreserved at -85 °C. These cells exhibited higher proliferative potential in vitro and are currently at passage 27.
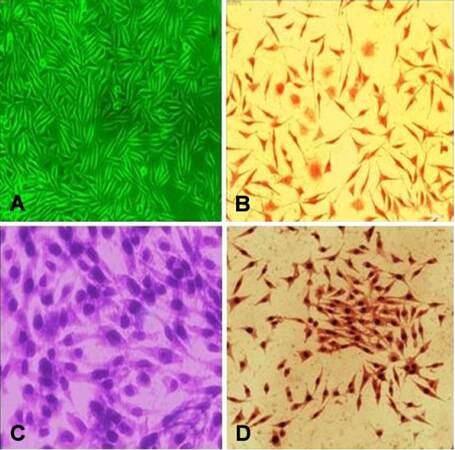
Figure 1. Phenotypic characterization of primary culture of CAFs isolated from metastatic CRC patient (×20). (A) CAFs isolated from the CRC patient cultured in vitro and exhibit spindle shaped morphology. Light microscopy of cultured CAFs for (B) Oil Red, (C) Giemsa stain and (D) Alzarin. CAFs: cancer associated fibroblasts; CRC: colorectal cancer
These CAF cells have a doubling time of 24 ± 0.7 h [Figure 2] and show an increased proliferative potential. Phase contrast microscopy study revealed that the cultured fibroblasts maintained their fibroblastic phenotype even in the absence of carcinoma cells and have a spindle-shaped appearance [Figure 1]. Further light microscopy study confirmed that primary CAFs indeed acquired a uniform spindle-shaped appearance and formed parallel arrays at confluence [Figure 1]. The majority of the CAFs displayed positive expression of vimentin [Figure 3] which is a marker for myofibroblastic differentiation. Contaminations by epithelial cells were excluded by negative expression for the epithelial marker keratin 18 [Figure 3].
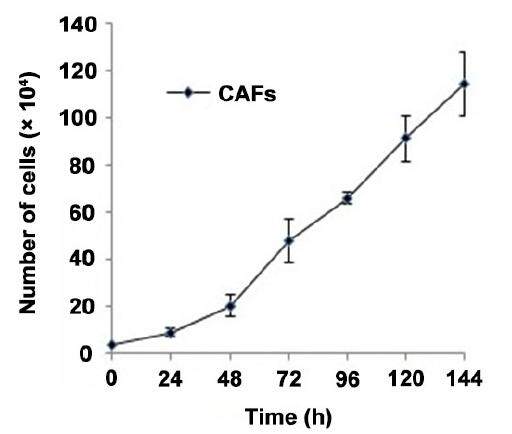
Figure 2. Cell kinetic of CAFs isolated from CRC patient. Nearly 40,000 of cells were seeded and cell count was done at different time interval. CAFs: cancer associated fibroblasts; CRC: colorectal cancer
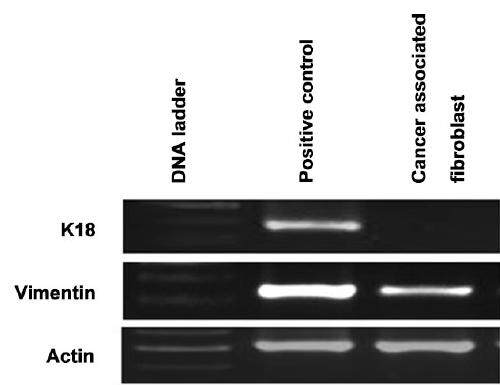
Figure 3. Gene expression of epithelial and myofibroblastic markers in CAFs isolated from CRC patient as measured by RT-PCR. CAFs: cancer associated fibroblasts; CRC: colorectal cancer
Isolated colorectal CAFs express the EMT inducible factors and pluripotency stem cell genes
CRC cells also undergo EMT which is associated with an increased stemness property and hence are able to regulate self-renewal because of certain transcription factors. The acquisition of cancer stem cell activity is a surrogacy for tumor-initiating cells and is believed to mediate disease relapse including poor prognosis in CRC.[9] We therefore evaluated the expression of EMT and pluripotency stem cells genes in stromal CAFs to investigate whether stromal cells acquire similar stem cell signature genes as observed. Qualitative RT-PCR has demonstrated that CAFs express high levels of SDF-1 and IL-6 [Figure 4] which has been shown to promote EMT in various carcinoma cells. These results suggest that the factors secreted by CAFs could induce the EMT phenotype in CRC. Besides, the stemness associated genes Oct4, Nanog and Sox2 were also expressed prominently in CAFs [Figure 5]. On the contrary, the level of Klf4 was quite low and it may indicate that the expression of Klf4 is inversely proportional to disease progression [Figure 5].
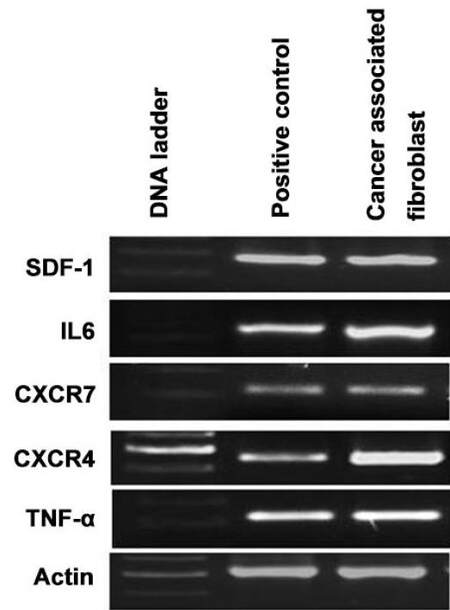
Figure 4. Gene expression of EMT inducer (SDF-1, IL6), chemokines and cytokines isolated in CAFs isolated from CRC patient as measured by RT-PCR. EMT: epithelial to mesenchymal transition; CAFs: cancer associated fibroblasts; CRC: colorectal cancer
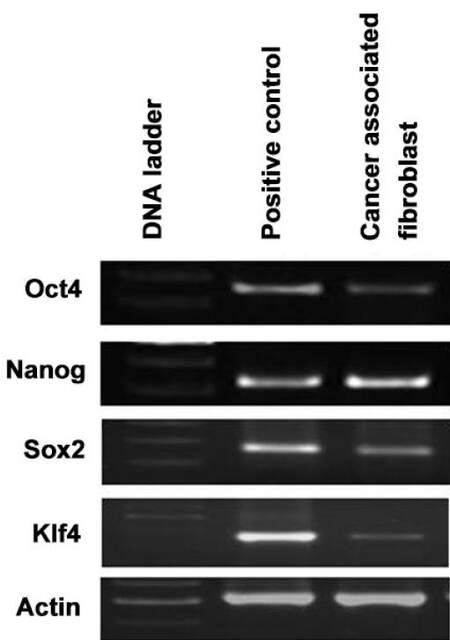
Figure 5. Gene expression of pluripotency stem markers in CAFs isolated from CRC patient as measured by RT-PCR. CAFs: cancer associated fibroblasts; CRC: colorectal cancer
CAFs exhibit expression of genes involved in cancer metastasis and tumorigenicity
Earlier studies have reported that CRCs possess a high metastatic potential with a concomitant ability to initiate tumor formation at secondary sites.[10] It is assumed that the acquisition of EMT phenotype allows CRCs to become more aggressive by enabling tissue invasiveness and increase in metastatic potential. Considering the dominant nature of stromal cells in dictating the fate of carcinoma cells, it is most likely that the genes expressed by stromal cells may also dynamically influence the metastatic potential of CRCs. We have shown that stromal CAFs indeed express matrix metalloproteinase genes such as MMP1, MMP2, and MMP9 [Figure 6].
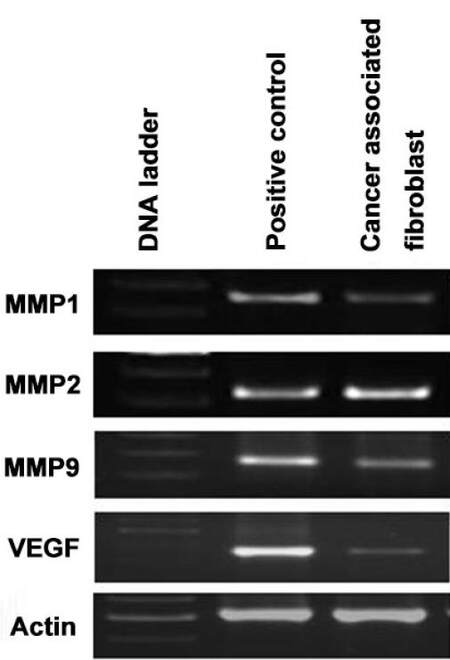
Figure 6. Gene expression of metastatic genes in CAFs isolated from CRC patient as measured by RT-PCR. CAFs: cancer associated fibroblasts; CRC: colorectal cancer
The inflammatory cytokine, TNF-α, which is directly involved in tumor invasion and metastasis, is also expressed by CAFs [Figure 4]. Another hallmark of tumor metastasis involves angiogenesis which is potentially mediated by vascular endothelial growth factor (VEGF). Interestingly, we have reported that CAFs express high levels of VEGF [Figure 6] and thus orchestrate metastatic progression of CRCs. In addition, we have shown that CAFs express chemokine receptors such as CXCR4 and CXCR7 which are involved in tumor progression [Figure 4]. Thus, the isolated CAFs in our study expressed genes that play a significant role in tumor metastasis.
Discussion
This study for the first time reported gene expression profiling of primary stromal fibroblast cells isolated from a patient with metastatic CRC. Although the stromal compartment in CRCs contains heterogeneity of cells,[11] we have performed prior separation of stromal cells from tumor cells and explored the gene profiling only on RNA isolated from stromal primary cultured fibroblasts. Interestingly, we have reported that isolated CAFs maintained their tumor-promoting phenotype and genotype in vitro even in absence of carcinoma cells which is suggestive of hereditable changes in CAFs.[10,11] However, CAFs show an increase in the proliferation capacity from passage 18 thereby indicating independence from the in-vivo tumor microenvironment for growth signals.
Another significant finding from this study is the molecular attributes of pluripotent gene expression which may be an inherent characteristic of stromal cells. A previous study has reported that the transcriptome of CRCs is also enriched with Oct4, Sox2, Nanog, and Klf4.[12] These cells, also termed cancer-initiating cells, represent a highly aggressive CRC which plays a significant role in chemoresistance and has been associated with poor prognosis.[13,14] We have reported that CAFs expressed similar pluripotent genes (Oct4, Sox2, Nanog, and Klf4) which suggests that both stromal cells and CRCs exhibit similar cancer stem cell gene expression and thus hypothesized that stromal cells may also regulate the cancer-stem cell properties in CRCs.
EMT represents one of the major events in CRCs and has been associated with high aggressiveness of the carcinoma with a concomitant increase in metastatic potential.[15-17] The inflammatory cytokine IL6 has been previously reported to be involved in expanding the cancer stem cell populations as well as inducing EMT. Both of these have implications in tumor metastasis and resistance to chemotherapeutic drugs.[18,19] Our result is in agreement with this study which has also reported that IL6 secreted by CAFs is a potent inducer of an EMT phenotype in breast cancer characterized by loss of e-cadherin expression and robust induction of a vimentin gene with increased proliferative indices.[20] Robust evidence has reported the role of chemokines in cancer.[21] In our study, we have shown that SDF-1 and CXCR4 are prominently expressed by the CAFs. There are reports which suggest that radiation could also induce EMT[22] and therefore we suggest that CAFs could be associated with tumor-regrowth in CRC after pre-operative chemoradiotherapy. A study by Onoue et al.[23] has demonstrated that EMT transition is also induced in oral squamous cell carcinoma by the SDF-1/CXCR4 axis. We believe that expression of the SDF-1 gene, as reported in our study, can also strongly contribute to EMT. An elevated level of another SDF-1 receptor, CXCR7, has been reported in CRC brain metastasis as well as in certain classes of aggressive colon tumor.[24] Thus, the expression of CXCR7 in our study could reflect the metastatic status of the CRC patient.
We have also shown that CAFs express MMP1, MMP2, and MMP9, which are proteases involved in tumor invasion and metastasis. Specifically, the collagenase MMP1 is a key mediator of primary tumor invasion and the use of neutralizing antibodies against MMP1 completely abolished colon cancer invasion.[25] Moreover, the polymorphism in the MMP1 gene has been associated with increased susceptibility to CRCs.[26] Thus, MMP1 secreted by the CAFs can act as a circulating tumor cell attractant for primary invasion of CRCs. The expression of gelatinases MMP2 and MMP9 has been associated with the worst outcome in many subsets of CRCs patients.[27] Angiogenesis is one of the hallmarks of metastatic cancer and is essential for the spread of CRCs. Clinical studies have reported that VEGF is the predominant proangiogenic factor in CRC and is overexpressed in 50% of CRCs, associated with distant metastasis.[28] We have reported similar expression of VEGF in the stromal CAFs from a CRC patient. Although mRNA of MMP1, MMP2, MMP9, and VEGF was confirmed in this study, more investigations are warranted in order to isolate and confirm proteins from the same genes.
Tumor necrosis factor alpha (TNF-α) plays a role in tumor growth by upregulating colony stimulating factor one (CSF-1) which in turn regulates the secretion of VEGF. Silencing of TNF-α in colon cancer cells with small-interfering RNAs inhibits the expression of CSF-1, associated with a decrease in the expression of VEGF and MMP2 mRNAs.[29] Our study has demonstrated similar expression of TNF-α with a concomitant expression of VEGF and MMP2 by stromal CAFs. Thus, the results clearly support and are in agreement with previous observations that CAFs indeed orchestrate the metastatic program required for primary tumor invasion and spread of cancer at secondary sites. More investigation should be undertaken in order to evaluate the positive expression of Alfa-SMA and FAP and negative expression of CK and CD45 that are considered as markers for CAFs in CRC. In addition, other experiments should be conducted that include the evaluation of CAFs, such as multiple colon CAF-derived factors that sustain proliferative signaling in CRC cells and support the cancer cells to resist cell death and evade growth suppressors including EGF, hepatocyte growth factor (HGF), IGF1/2, PGE-2, PDGF, fibroblast growth factor (FGF)-1, and VEGF.
In conclusion, the present study is the first report of the isolation of CAFs from PBMCs of metastatic CRC. These were fast growing cells and showed all the characteristics of metastatic tumor cells. Molecular profiling of these cells prominently expressed metastatic genes such as MMP1, MMP2, MMP9, and VEGF, which have promoted the metastatic process in CRC. Our study therefore hypothesizes that the targeting of these genes and pathways responsible for the metastatic process probably helps in reducing the metastatic potencies of CAF stromal cells which may allow clinicians to control the spreading of these cells at distant organs and to create better therapeutic solutions for colon cancer patients.
Acknowledgments
The authors wish to thank the management of Jaslok Hospital and Research Center for allowing them to carry out this work at their institute and for their constant support.
Financial support and sponsorship
Nil.
Conflicts of Interest
There are no conflicts of interest.
Patient consent
Consent was obtained from the patient involved in the study.
Ethics approval
Ethics approval was obtained prior to the commencement of the study.
REFERENCES
1. Aune D, Chan DS, Lau R, Vieira R, Greenwood DC, Kampman E, Norat T. Dietary fibre, whole grains, and risk of colorectal cancer: systematic review and dose-response meta-analysis of prospective studies. BMJ 2011;343:d6617.
2. Conti J, Thomas G. The role of tumour stroma in colorectal cancer invasion and metastasis. Cancers (Basel) 2011;3:2160-8.
3. Guillem JG, Chessin DB, Cohen AM, Shia J, Mazumdar M, Enker W, Paty PB, Weiser MR, Klimstra D, Saltz L, Minsky BD, Wong WD. Long-term oncologic outcome following preoperative combined modality therapy and total mesorectal excision of locally advanced rectal cancer. Ann Surg 2005;241:829-36.
4. Calon A, Lonardo E, Berenguer-Llergo A, Espinet E, Hernando-Momblona X, Iglesias M, Sevillano M, Palomo-Ponce S, Tauriello DV, Byrom D, Cortina C, Morral C, Barceló C, Tosi S, Riera A, Attolini CS, Rossell D, Sancho E, Batlle E. Stromal gene expression defines poor-prognosis subtypes in colorectal cancer. Nat Genet 2015;47:320-9.
5. Farmer P, Bonnefoi H, Anderle P, Cameron D, Wirapati P, Becette V, André S, Piccart M, Campone M, Brain E, Macgrogan G, Petit T, Jassem J, Bibeau F, Blot E, Bogaerts J, Aguet M, Bergh J, Iggo R, Delorenzi M. A stroma-related gene signature predicts resistance to neoadjuvant chemotherapy in breast cancer. Nat Med 2009;15:68-74.
6. Isella C, Terrasi A, Bellomo SE, Petti C, Galatola G, Muratore A, Mellano A, Senetta R, Cassenti A, Sonetto C, Inghirami G, Trusolino L, Fekete Z, De Ridder M, Cassoni P, Storme G, Bertotti A, Medico E. Stromal contribution to the colorectal cancer transcriptome. Nat Genet 2015;47:312-9.
7. Potdar PD, Shetti AU. Evaluation of anti-metastatic effect of chitosan nanoparticles on esophageal cancer-associated fibroblasts. J Cancer Metastatis Treat 2016;2:259-67.
8. Potdar P, Sutar J. Establishment and molecular characterization of mesenchymal stem cell lines derived from human visceral & subcutaneous adipose tissues. J Stem Cells Regen Med 2010;6:26-35.
9. Ricci-Vitiani L, Lombardi DG, Pilozzi E, Biffoni M, Todaro M, Peschle C, De Maria R. Identification and expansion of human colon-cancer-initiating cells. Nature 2007;445:111-5.
10. Tommelein J, Verset L, Boterberg T, Demetter P, Bracke M, De Wever O. Cancer-associated fibroblasts connect metastasis-promoting communication in colorectal cancer. Front Oncol 2015;5:63.
11. Linnekamp JF, Wang X, Medema JP, Vermeulen L. Colorectal cancer heterogeneity and targeted therapy: a case for molecular disease subtypes. Cancer Res 2015;75:245-9.
12. Saiki Y, Ishimaru S, Mimori K, Takatsuno Y, Nagahara M, Ishii H, Yamada K, Mori M. Comprehensive analysis of the clinical significance of inducing pluripotent stemness-related gene expression in colorectal cancer cells. Ann Surg Oncol 2009;16:2638-44.
13. Orimo A, Gupta PB, Sgroi DC, Arenzana-Seisdedos F, Delaunay T, Naeem R, Carey VJ, Richardson AL, Weinberg RA. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 2005;121:335-48.
14. Ishii G, Hashimoto H, Asada K, Ito T, Hoshino A, Fujii S, Kojima M, Kuwata T, Harigaya K, Nagai K, Ushijima T, Ochiai A. Fibroblasts associated with cancer cells keep enhanced migration activity after separation from cancer cells: a novel character of tumor educated fibroblasts. Int J Oncol 2010;37:317-25.
15. Haraguchi N, Utsunomiya T, Inoue H, Tanaka F, Mimori K, Barnard GF, Mori M. Characterization of a side population of cancer cells from human gastrointestinal system. Stem Cells 2006;24:506-13.
16. Ieta K, Tanaka F, Haraguchi N, Kita Y, Sakashita H, Mimori K, Matsumoto T, Inoue H, Kuwano H, Mori M. Biological and genetic characteristics of tumor-initiating cells in colon cancer. Ann Surg Oncol 2008;15:638-48.
17. De Sousa E Melo F, Wang X, Jansen M, Fessler E, Trinh A, de Rooij LP, de Jong JH, de Boer OJ, van Leersum R, Bijlsma MF, Rodermond H, van der Heijden M, van Noesel CJ, Tuynman JB, Dekker E, Markowetz F, Medema JP, Vermeulen L. Poor-prognosis colon cancer is defined by a molecularly distinct subtype and develops from serrated precursor lesions. Nat Med 2013;19:614-8.
18. Iliopoulos D, Hirsch HA, Wang G, Struhl K. Inducible formation of breast cancer stem cells and their dynamic equilibrium with non-stem cancer cells via IL6 secretion. Proc Natl Acad Sci U S A 2011;108:1397-402.
19. Korkaya H, Kim GI, Davis A, Malik F, Henry NL, Ithimakin S, Quraishi AA, Tawakkol N, D'Angelo R, Paulson AK, Chung S, Luther T, Paholak HJ, Liu S, Hassan KA, Zen Q, Clouthier SG, Wicha MS. Activation of an IL6 Inflammatory Loop Mediates Trastuzumab Resistance in HER2+ breast cancer by expanding the cancer stem cell population. Mol Cell 2012;47:570-84.
20. Sullivan NJ, Sasser AK, Axel AE, Vesuna F, Raman V, Ramirez N, Oberyszyn TM, Hall BM. Interleukin-6 induces an epithelial-mesenchymal transition phenotype in human breast cancer cells. Oncogene 2009;28:2940-7.
21. Verbeke H, Struyf S, Laureys G, Van Damme J. The expression and role of CXC chemokines in colorectal cancer. Cytokine Growth Factor Rev 2011;22:345-58.
22. Tsukamoto H, Shibata K, Kajiyama H, Terauchi M, Nawa A, Kikkawa F. Irradiation-induced epithelial-mesenchymal transition (EMT) related to invasive potential in endometrial carcinoma cells. Gynecol Oncol 2007;107:500-4.
23. Onoue T, Uchida D, Begum NM, Tomizuka Y, Yoshida H, Sato M. Epithelial-mesenchymal transition induced by the stromal cell-derived factor-1/CXCR4 system in oral squamous cell carcinoma cells. Int J Oncol 2006;29:1133-8.
24. Heckmann D, Maier P, Laufs S, Li L, Sleeman JP, Trunk MJ, Leupold JH, Wenz F, Zeller WJ, Fruehauf S, Allgayer H. The disparate twins: a comparative study of CXCR4 and CXCR7 in SDF-1α-induced gene expression, invasion and chemosensitivity of colon cancer. Clin Cancer Res 2014;20:604-16.
25. Raufman JP, Cheng K, Saxena N, Chahdi A, Belo A, Khurana S, Xie G. Muscarinic receptor agonists stimulate matrix metalloproteinase 1-dependent invasion of human colon cancer cells. Biochem Biophys Res Commun 2011;415:319-24.
26. Decock J, Paridaens R, Ye S. Genetic polymorphisms of matrix metalloproteinases in lung, breast and colorectal cancer. Clin Genet 2008;73:197-211.
27. Said AH, Raufman JP, Xie G. The role of matrix metalloproteinases in colorectal cancer. Cancers (Basel) 2014;6:366-75.
28. Araújo RF Jr, Lira GA, Vilaça JA, Guedes HG, Leitão MC, Lucena HF, Ramos CC. Prognostic and diagnostic implications of MMP-2, MMP-9, and VEGF-α expressions in colorectal cancer. Pathol Res Pract 2015;211:71-7.
Cite This Article
Export citation file: BibTeX | RIS
OAE Style
Potdar PD, Chaudhary S. Molecular characterization of cancer-associated fibroblasts isolated from human colorectal cancer as a major stromal cell component promoting metastasis. J Unexplored Med Data 2017;2:1-8. http://dx.doi.org/10.20517/2572-8180.2016.10
AMA Style
Potdar PD, Chaudhary S. Molecular characterization of cancer-associated fibroblasts isolated from human colorectal cancer as a major stromal cell component promoting metastasis. Journal of Unexplored Medical Data. 2017; 2: 1-8. http://dx.doi.org/10.20517/2572-8180.2016.10
Chicago/Turabian Style
Potdar, Pravin D., Shahid Chaudhary. 2017. "Molecular characterization of cancer-associated fibroblasts isolated from human colorectal cancer as a major stromal cell component promoting metastasis" Journal of Unexplored Medical Data. 2: 1-8. http://dx.doi.org/10.20517/2572-8180.2016.10
ACS Style
Potdar, PD.; Chaudhary S. Molecular characterization of cancer-associated fibroblasts isolated from human colorectal cancer as a major stromal cell component promoting metastasis. J. Unexplored. Med. Data. 2017, 2, 1-8. http://dx.doi.org/10.20517/2572-8180.2016.10
About This Article
Copyright

Author Biographies

Data & Comments
Data


 Cite This Article 7 clicks
Cite This Article 7 clicks

 Like This Article 30
likes
Like This Article 30
likes






Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.